Our research interests are focused on developing and applying
solid-state NMR methods to determine structural constraints in
biologically relevant systems that are difficult to fully
characterize by other means in order to contribute to our
overall understanding of biological processes.
In the near future, we will be working on determining the
internal structure of the β2 microglobulin protein in
its fibrillar form.
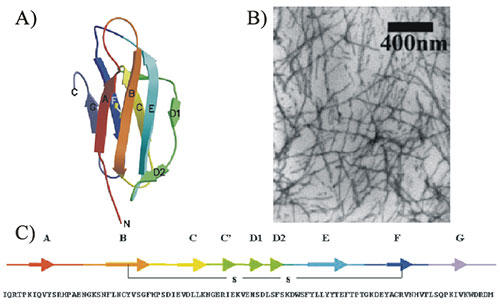
Figure 1: β2
microglobulin. A) Native fold. B) Electron micrograph of β2
microglobulin amyloid fibrils. C) Amino acid sequence.
(Images from Jones et al., J. Mol. Biol. (2003) 325
pp249-257).
Filamentous fibrillar amyloid aggregates are interesting to
study because of the many diseases with which they are
associated, but also from the standpoint of understanding
protein folding. A large number of disparate proteins have been
shown to form fibrils with morphologies similar to that found in
Fig. 1B. Amyloid fibrils exhibit a characteristic ‘cross-β’
scattering pattern from X-ray fiber diffraction measurements
indicating the presence of β-sheets oriented perpendicular to
the long axis of the fibril. While the internal structures of at
least a few of these different fibrils have been shown to be
different in detail, it is unknown why the overall morphologies
are so similar. Considerable effort is being expended in
elucidating the intermolecular interactions that stabilize
amyloid formation, the mechanisms of amyloid formation from
peptide monomers or oligomers, and the internal molecular
structure of peptides in amyloid fibrils.
We will be focusing on the β2 microglobulin protein
(Fig. 1). β2 microglobulin is a relatively small
protein with 99 residues that is the major component of dialysis
related amyloid. Dialysis related amyloidosis is a serious
complication for patients receiving long term hemodialysis,
resulting in carpal tunnel syndrome and/or arthritis-like pain
in joints and tendons. Eventually, a majority of long-term
hemodialysis patients experience the debilitating effects of the
disease due to a buildup of fibrillar amyloid material in bone
and joint tissues. Because β2 microglobulin is a
small protein and readily expressed in a variety of expression
systems, it has been used extensively in amyloid formation
studies, but little is known about its internal molecular
structure in amyloid fibrils beyond the fact that it contains
the ‘cross-β’ motif common to all amyloid fibrils. Since amyloid
fibrils are ill suited to study by solution-state NMR because
they are insoluble aggregates, and neither is the detailed
internal structure of fibrils obtainable by crystallography
because fibrils do not exhibit the long range crystalline order
required, we propose to apply a set of solid-state NMR structure
elucidation tools to answer questions about the structure of the
β2 microglobulin protein in its fibrillar form.
Figure 2:
Experimental measurements to determine the global fold of
the peptide in the amyloid fibril. A) Multiple quantum
experiments to distinguish between different organizational
structures. B) Backbone torsion angle measurements at
particular locations. C) Proximity measurements between
different parts of the peptide.
Dipolar couplings, chemical shift anisotropies (CSAs), and
their cross correlations can be measured at particular locations
in the peptide to give structural constraints. For instance,
distances up to ~5-6 Å between two isotopic labels in a peptide
(Fig. 2C) can be determined either by homonuclear or
heteronuclear dipolar recoupling experiments. Such distance
measurements can be used, for instance, to verify a hairpin, in
which a peptide folds back on itself (as is postulated for the
P72-M99 fragment). A variety of dipolar recoupling and
cross-correlation experiments measure one or both of the peptide
backbone torsion angles (Fig. 2B) at a single amino acid
position for a direct measure of the secondary structure at that
position. Additionally, solid-state NMR may be used to
distinguish between fibril models with different organization of
peptides internal to the fibril. In particular, so-called ‘spin
counting’ multiple quantum experiments can estimate the number
of isotopic labels within a dipolar-coupling network. Fig. 2A
shows a series of different internal organizations of peptides
in a fibril that differ in the number of isotopic labels that
are in close proximity to each other. Multiple quantum (MQ)
spectra of a parallel β-sheet organization would show high
orders of MQ coherence, while the anti-parallel case would show
no higher orders since none of the isotopic labels are in
proximity to each other. Dimeric and trimeric groupings would
correspondingly show either two or three quantum coherences at
most.